暘壔偟偨僯儏乕儘儞偑堏摦傪奐巒偡傞儊僇僯僘儉
敪惗婜偺戝擼怴旂幙偵偍偄偰枹暘壔側恄宱宯慜嬱嵶朎偐傜偼師乆偲弴彉傛偔條乆側僯儏乕儘儞偑嶻傒弌偝傟傞偑丄 偦偺帪娫弴彉偑旂幙斅撪偺嬻娫弴彉偵懳墳偡傞偙偲乮inside-out條幃乯偑傛偔抦傜傟偰偄傞丅 偙偺傛偆側弴彉偺懳墳傪惓妋偵峴側偄惓偟偄旂幙峔抸傪幚尰偡傞偨傔偵偼丄僯儏乕儘儞偑暘壔捈屻偵堏摦偡傞偙偲偑廳梫偱偁傞丅 偟偐偟丄乽僯儏乕儘儞塣柦寛掕偲堏摦奐巒偺僇僢僾儕儞僌乿傪惂屼偡傞儊僇僯僘儉偼晄柧偱偁偭偨丅
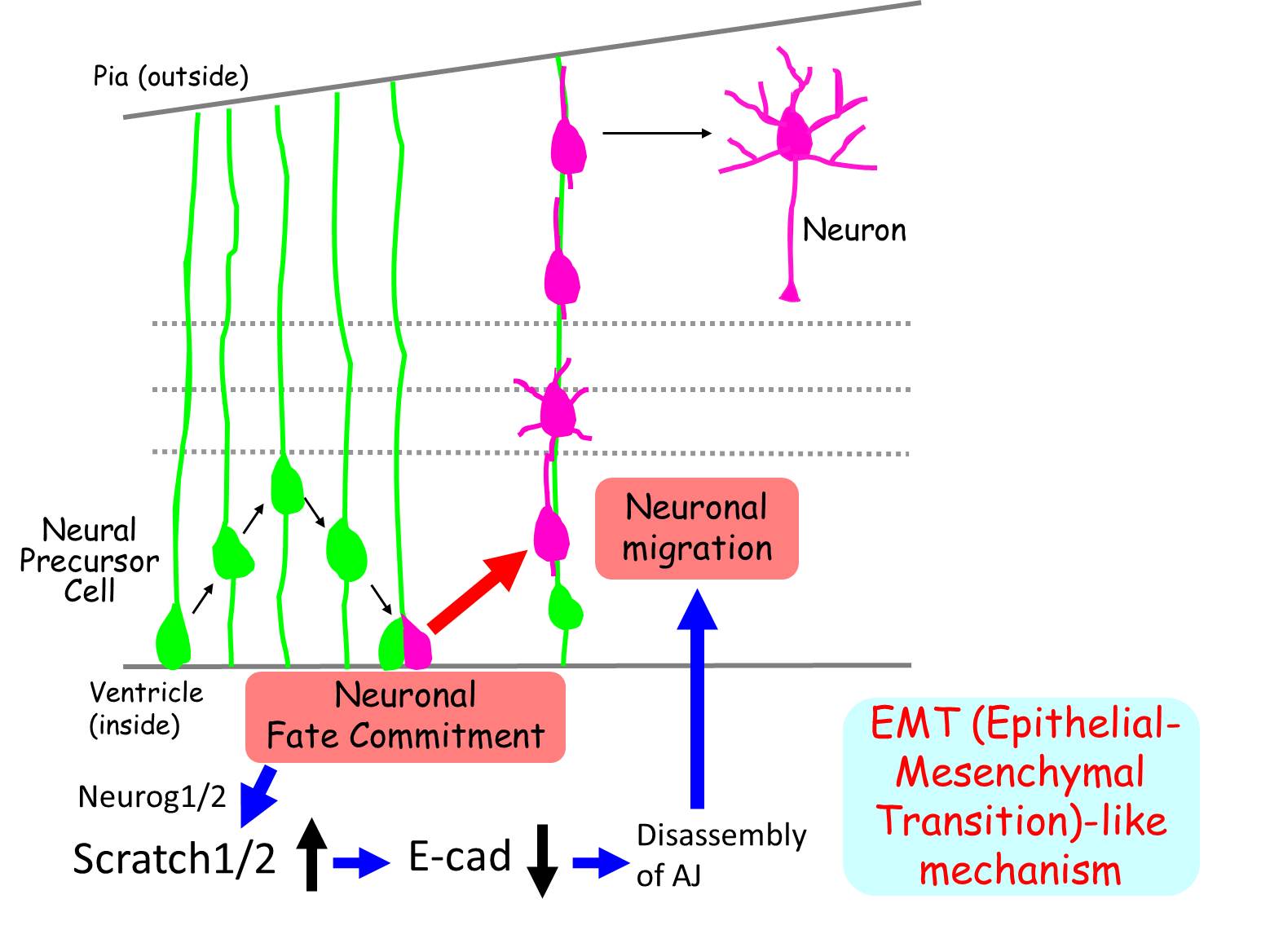
僯儏乕儘儞暘壔寛掕偵偼Neurog2側偳偺僾儘僯儏乕儔儖揮幨場巕偑廳梫側栶妱傪壥偨偡偙偲偑抦傜傟偰偄傞丅 杮尋媶偱偼丄偙傟傜偺僾儘僯儏乕儔儖揮幨場巕偺壓棳偱Snail superfamily揮幨場巕偱偁傞Scratch1/2偑敪尰偡傞偙偲傪尒弌偟偨丅 偝傜偵丄Scratch1/2偺夁忚敪尰偵傛偭偰嵶朎偑擼幒柺偐傜棧扙偟偰擼幒懷偺奜懁偵岦偐偭偰堏摦偡傞偙偲丄 媡偵Scratch1/2偺僲僢僋僟僂儞偵傛偭偰暘壔偟偨僯儏乕儘儞偺堏摦偺奐巒偑慾奞偝傟攝抲偑堎忢偵側傞偙偲傪尒弌偟偨丅 埲忋偺寢壥偐傜丄Scratch1/2偑僯儏乕儘儞傊偺暘壔寛掕屻偵堏摦奐巒傪桿摫偟偰偄傞偙偲偑帵嵈偝傟偨丅
恄宱宯慜嬱嵶朎偼丄嵶朎娫愙拝暘巕E-cadherin側偳傪嵶朎昞柺偵敪尰偟丄偍屳偄偵嵶朎娫愙拝傪宍惉偟偰偄傞丅 嫽枴怺偄偙偲偵丄Scratch1/2偼E-cadherin偺敪尰傪梷惂偟偰嵶朎娫愙拝傪掅壓偝偣傞偙偲偵傛傝擼幒柺偐傜偺棧扙傪桿摫偡傞偙偲偑柧傜偐偵側偭偨丅
杮尋媶偐傜丄Scratch偑僯儏乕儘儞偺塣柦寛掕偲堏摦奐巒偺僇僢僾儕儞僌偵昁恵偺暘巕偱偁傞偙偲偑帵嵈偝傟偨丅 Scratch偼戝擼怴旂幙埲奜偺拞悤恄宱宯偵偍偄偰傕僯儏乕儘儞暘壔偺僞僀儈儞僌偱暆峀偔敪尰偑娤嶡偝傟傞偙偲偐傜丄 変乆偑柧傜偐偵偟偨僯儏乕儘儞偺堏摦奐巒儊僇僯僘儉偼拞悤恄宱宯偵嫟捠偟偨傕偺偱偁傞壜擻惈偑峫偊傜傟傞丅
峏偵丄僯儏乕儘儞偺堏摦奐巒偺桿摫偼乽忋旂娫梩揮姺乿乮嵶朎摨巑偱偺愙拝偑嫮屌側嵶朎廤抍偺拞偐傜愙拝偑庛偄嵶朎偑嶻傑傟丄廤抍偐傜棧扙偡傞尰徾乯 偵椶帡偟偨儊僇僯僘儉偱惂屼偝傟偰偄傞偙偲偑帵嵈偝傟傞丗 側偤側傜丄?@乯Scratch偑忋旂娫梩揮姺偵廳梫側栶妱傪壥偨偡Snail superfamily偵懏偟丄 ?A乯Scratch偵傛傞堏摦奐巒惂屼儊僇僯僘儉偑揟宆揑側忋旂娫梩揮姺桿摫儊僇僯僘儉偵椶帡偟偰偄傞偐傜偱偁傞丅 忋旂娫梩揮姺偼屄懱偺敪惗偵廳梫側庬乆偺応柺偱梡偄傜傟傞尰徾偱偁傞偑丄偙傟傑偱擼敪惗偵娭傢傞偲偄偆徹嫆偼尒偮偐偭偰偄側偐偭偨丅 廬偭偰丄杮尋媶偼擼敪惗偵忋旂娫梩揮姺偑娭傢偭偰偄傞偙偲傪帵嵈偡傞弶傔偰偺惉壥偱偁傞偲峫偊傜傟傞 (Itoh et al., Nature Neuroscience, 2013)丅