ポリコーム群タンパク質複合体による神経幹細胞運命転換
発生時期依存的な神経幹細胞の運命転換が、「細胞自律的に何らかのタイマーを使って起こっているのか?」、 あるいは「細胞外からの指令を受けて起こっているのか?」という興味深い問題がある。 これまで、幹細胞をひとつだけ取り出してin vitroで培養しても運命転換が観察されることから、 少なくとも細胞自律的なメカニズムが存在するだろうと考えられてきた(細胞外因子の寄与を否定するものではないが)。 我々は、この細胞自律的なメカニズムの実態として、ポリコーム群タンパク質複合体によるクロマチン状態のエピジェネティックな変化が 貢献していることを見出した。以下にその概要を述べたい。
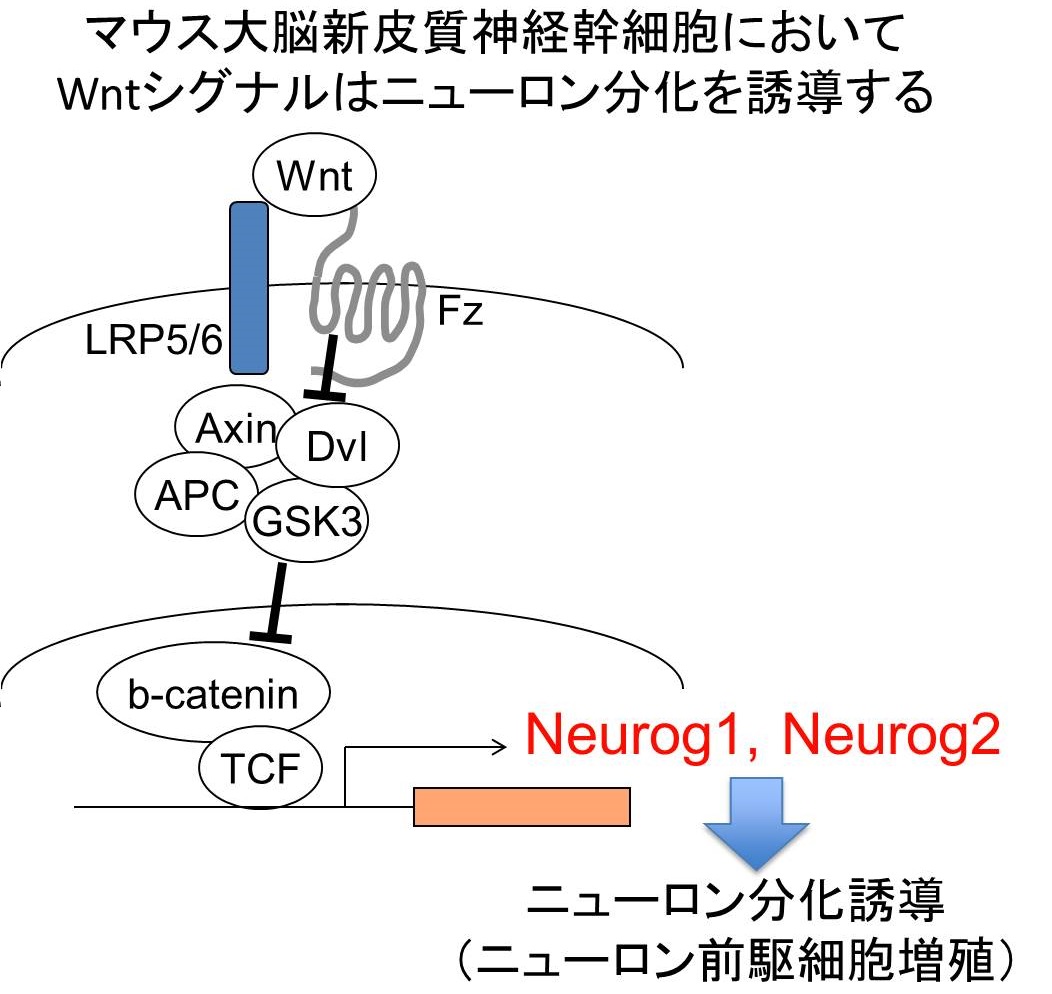
当研究室では以前に、マウス胎児大脳新皮質由来神経幹細胞のニューロン分化をinstructiveに誘導するシグナル伝達として Wnt-βcatenin経路が重要である事を報告した (Hirabayashi et al. Development 2004)。 Wnt-βcatenin経路はニューロン分化運命の決定因子であるNeurog1, Neurog2の発現を誘導し、分化を惹起する。
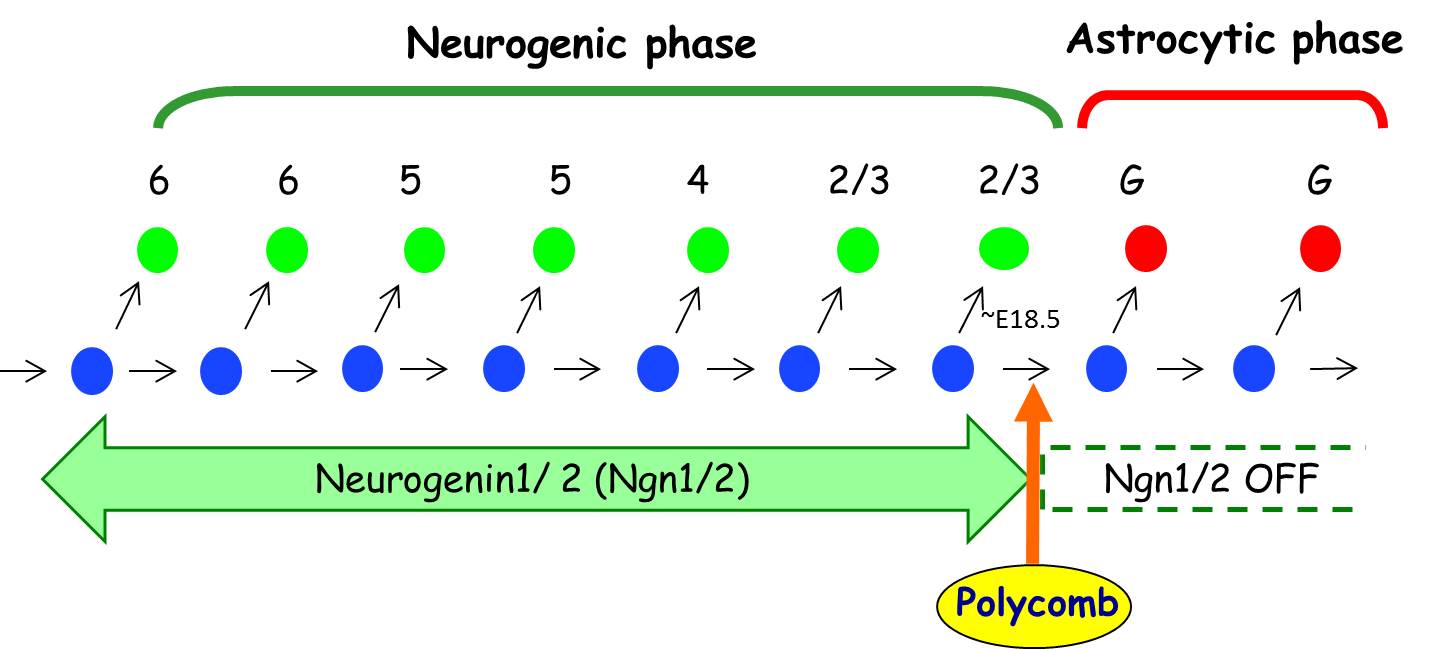
なぜ大脳発生において、ニューロン分化期が特定の時期に限定されているのか?という問いに対して、 もしWnt-βcatenin経路が特定の時期にのみ活性化していれば説明がつくかもしれない。しかしながら、神経幹細胞においてWnt-βcatenin経路は (ニューロン分化がほぼ起こらない)グリア分化期においても活性化していた。しかし転写因子βcateninの直接のターゲットであるNeurog1の転写は 誘導されていない。なぜか。
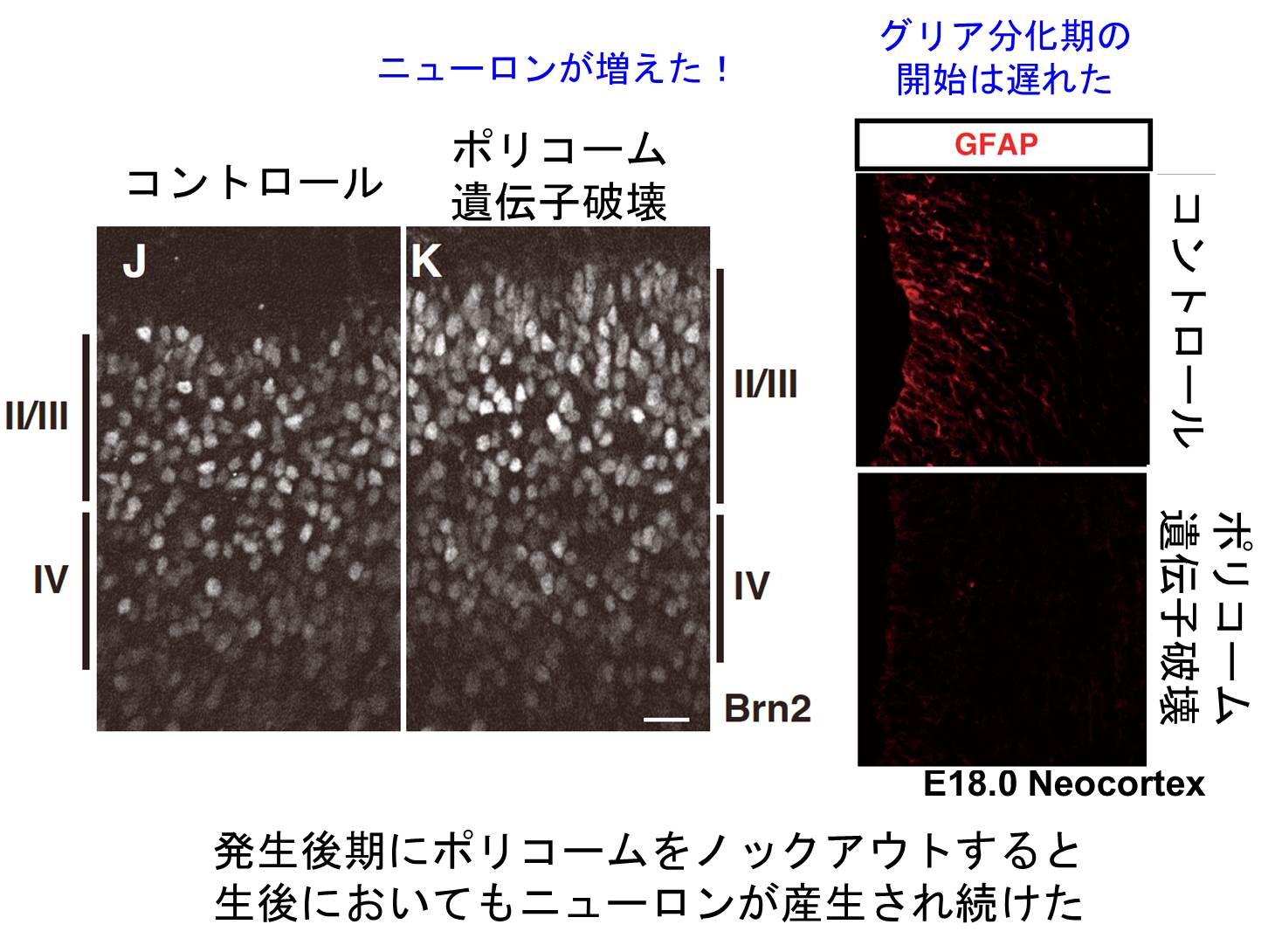
我々は、実はグリア分化期になると、Neurog1遺伝子座がポリコーム群タンパク質複合体(PcG)により「不活性型のクロマチン状態」になっていること、 それが故にWntシグナルが入ってもNeurog1遺伝子の転写が誘導されないこと、を見出した (Hirabayashi et al. Neuron 2009)。 PcGの必須構成因子Ring1Bをグリア分化期に入る前に神経幹細胞で欠損させると、Neurog1遺伝子座が許容状態になり、 普段ニューロン分化が起こらなくなる時期になってもニューロンを作り続けた。
PcGはNeurog1に限らず、時期依存的な運命転換に関わる多くの遺伝子座を、特定の時期に不活性化することが明らかになってきた(未発表)。 従って、PcGこそが細胞自律的なタイマーに従って運命転換を司るマシナリーの中心因子であることが示唆された。