摉尋媶幒偼丄摦暔嵶朎丄摿偵恄宱嵶朎傪嵽椏偵丄嵶朎偺塣柦惂屼儊僇僯僘儉 傪暘巕儗儀儖偱棟夝偡傞偙偲傪栚巜偟偰偄傞丅
1. 偼偠傔偵
2. 嵶朎偺惗巰媦傃塣摦惈偺惂屼
3. 恄宱宯慜嬱嵶朎偺塣柦惂屼
1.偼偠傔偵
惗暔偺敪惗夁掱偵偍偄偰丄嵶朎偼憹怋偲惗巰傪尩枾偵惂屼偝傟側偑傜丄條乆側宍幙傪帩偭偨嵶朎傊偲暘壔偟偰偄偔丅傑偨敪惗屻偺惉懱偵偍偄偰傕丄屄乆偺嵶朎偺憹怋丒暘壔丒嵶朎巰偼丄屄懱偲偟偰偺峆忢惈傪曐偮偨傔偵尩枾偵惂屼偝傟偰偄傞丅偙偺屄乆偺嵶朎塣柦偼丄婎杮揑偵嵶朎奜僔僌僫儖偑嵶朎枌忋偺庴梕懱傪夘偟偰僐儞僩儘乕儖偟偰偄傞偙偲偑懡偄偑丄変乆偼丄庴梕懱偵巒傑傞乽嵶朎撪僔僌僫儖揱払乿偑偄偐偵忣曬傪妀偵揱偊丄堚揱忣曬偺惂屼偵寢傃偮偗偰偄傞偺偐丄偵拲栚偟偰偄傞丅変乆偺僌儖乕僾偼丄乽嵶朎撪僔僌僫儖揱払乿偵偍偄偰拞怱揑側栶妱傪壥偨偟偰偄傞
MAP僉僫乕僛偺僇僗働乕僪 偺摨掕偵峷專偟丄偦偺惂屼婡峔傪柧傜偐偵偟偰偒偨丅MAP僉僫乕僛僇僗働乕僪偼丄尨娻堚揱巕Ras偺壓棳偱妶惈壔偟嵶朎憹怋僔僌僫儖偲偟偰摥偔偩偗偱側偔丄條乆側暘壔偺僔僌僫儖揱払偵偍偄偰傕廳梫側栶妱傪壥偨偟偰偄傞丅MAP僉僫乕僛僇僗働乕僪偑愐捙摦暔偩偗偱側偔丄峺曣丒怉暔丒柍愐捙摦暔側偳峀斖側惗暔庬偱嫟捠偵梡偄傜傟偰偄傞乽僔僌僫儖揱払儌僕儏乕儖乿偱偁傞偙偲傕暘偐偭偰偒偨丅傑偨丄MAP僉僫乕僛偺僗乕僷乕僼傽儈儕乕乮JNK,
p38側偳乯偺拞偵偼丄僗僩儗僗偵墳摎偟偰傾億僩乕僔僗乮嵶朎偺帺嶦乯偵娭傢傞傕偺偑偁傞偙偲傕帵偟偰偒偨丅偟偐偟側偑傜丄屄乆偺尰徾傪堷偒婲偙偡偲偒偺MAP僉僫乕僛偺僞乕僎僢僩偵偮偄偰偼枹偩昁偢偟傕柧傜偐偱偼側偄丅変乆偼摿偵嵶朎偺惗巰偲嵶朎娻壔偵偍偗傞僔僌僫儖揱払偵拲栚偟偰夝愅傪峴偭偰偄傞丅
2.丂嵶朎偺惗巰媦傃塣摦惈偺惂屼
嵶朎偺惗巰偺惂屼偑偆傑偔偄偐側偗傟偽丄昁梫側嵶朎偑巰傫偩傝丄巰偸傋偒嵶朎乮椺偊偽DNA懝彎傪庴偗偨嵶朎丄僂僀儖僗偵姶愼偟偨嵶朎傗僈儞嵶朎乯偑巰側側偔側偭偰丄屄懱偺惗柦傪嫼偐偡丅嵶朎偺惗巰偼丄
乽惗懚僔僌僫儖揱払乿 偲乽巰僔僌僫儖揱払乿偺僶儔儞僗偵傛偭偰岻柇偵惂屼偝傟偰偄傞偑丄変乆偼偙偺儊僇僯僘儉偺夝柧偵庢傝慻傫偱偄傞丅
慄拵偐傜歁擕椶傑偱恑壔揑偵曐懚偝傟偰偄傞傾億僩乕僔僗偺僐傾僾儘僌儔儉偼丄僇僗儁乕僗偲屇偽傟傞僾儘僥傾乕僛傪拞怱偲偟偰偍傝丄僞儞僷僋幙暘夝傪夘偟偰乽晄壜媡揑偵乿嵶朎巰傪桿摫偡傞丅慄拵偺僾儘僌儔儉嵶朎巰偱偼丄懱嵶朎偺偆偪偳偺嵶朎偑巰偸偐偑偁傜偐偠傔寛掕偟偰偄傞丅堦曽歁擕椶偱偼丄廃埻偺娐嫬傗庴偗偨僟儊乕僕偺掱搙傪憤崌揑偵敾抐偟丄嵶朎偑巰偸偐斲偐傪寛傔傞偨傔偺乽壜媡揑僾儘僙僗乿偑懚嵼偟丄晄壜媡揑側僇僗儁乕僗宱楬傪惂屼偡傞偲峫偊傜傟傞丅変乆偼丄嵶朎偺惗巰寛掕偵娭傢傞丄儕儞巁壔傪戙昞偲偟偨乽壜媡揑僾儘僙僗乿偵拲栚偟丄尰嵼埲壓偺揰傪専摙偟偰偄傞丅
(1) Akt偵傛傞惗懚懀恑儊僇僯僘儉偲娻壔傊偺峷專
Akt 偼懡偔偺宯偱惗懚懀恑偵拞怱揑側栶妱傪壥偨偡偙偲偑帵偝傟偰偄傞丅Akt偼僙儕儞/僗儗僆僯儞僉僫乕僛偱偁傞偑丄婎幙偵娭偟偰偼婔偮偐岓曗偑偁傞傕偺偺枹偩Akt偵傛傞惗懚懀恑儊僇僯僘儉偺慡梕偼柧傜偐偱偼側偄丅変乆偼丄Akt偺嶌梡揰偲偟偰丄
恾偵帵偟偨暋悢偺僞乕僎僢僩傪柧傜偐偵偟偮偮偁傞丅偙偺傛偆偵Akt偼丄傾億僩乕僔僗僔僌僫儖揱払偺暋悢偺僗僥僢僾傪摨帪偵梷惂偡傞偙偲偱妋幚偵惗懚傪曐徹偟偰偄傞傕偺偲峫偊傜傟傞丅
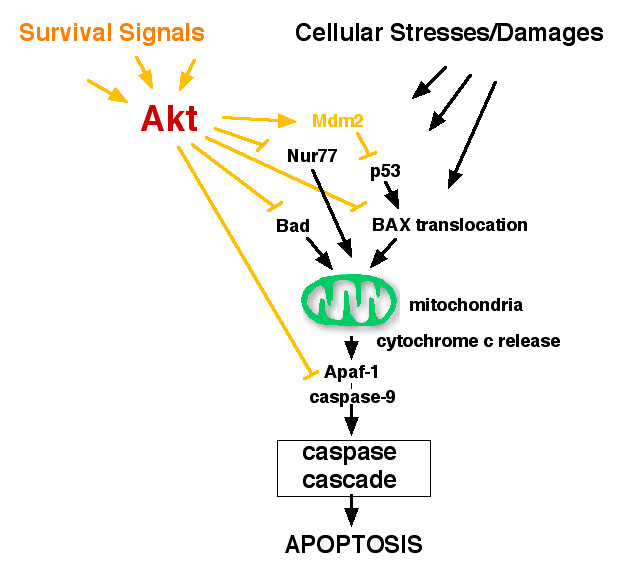
Akt偼尨娻堚揱巕偲偟偰摨掕偝傟丄幚嵺偵懡偔偺娻慻怐偱Akt偺妶惈壔偑尒傜傟偰偍傝丄娻壔偲偺怺偄娭楢偑帵嵈偝傟偰偄傞丅堦斒揑偵娻壔懀恑婡峔偲偟偰丄1.
憹怋懀恑丄2. 惗懚懀恑丄3.塣摦惈丒 怹弫惈偺忋徃丄4. 寣娗怴惗丄5. 晄巰壔丄側偳偑嫇偘傜傟傞偑丄Akt偑偳偺梫場偱娻壔偵峷專偟偰偄傞偐昁偢偟傕柧傜偐偱偼側偄丅変乆偼丄忋婰偺惗懚懀恑偩偗偱側偔丄
塣摦惈偺忋徃 側偳丄Akt偵傛傞娻壔偺儊僇僯僘儉偵娭偟偰暿偺妏搙偐傜傕専摙偟偰偄傞丅
(2) JNK埶懚揑側嵶朎巰桿摫儊僇僯僘儉偲偦偺惗棟揑堄媊
JNK偼條乆側僗僩儗僗巋寖偱嫟捠偵妶惈壔偡傞僙儕儞/僗儗僆僯儞僉僫乕僛偱偁傞丅変乆傪娷傔偨婔偮偐偺僌儖乕僾偼丄JNK宱楬偑僗僩儗僗偵傛傞僇僗儁乕僗宱楬偺妶惈壔偵廳梫側栶妱傪壥偨偟偰偄傞偙偲傪帵偟偰偒偨丅偝傜偵変乆偼丄
JNK宱楬偺妶惈壔偑丄僇僗儁乕僗旕埶懚揑嵶朎巰傪桿摫偟偆傞 偙偲傪帵偟偨丅偦偙偱尰嵼丄JNK偺(i)僇僗儁乕僗妶惈壔偵偍偗傞僞乕僎僢僩丄媦傃(ii)僇僗儁乕僗旕埶懚揑嵶朎巰偵偍偗傞僞乕僎僢僩丄傪専摙偟偰偄傞丅嬤擭丄條乆側敪惗夁掱傗昦棟揑嵶朎巰偺夁掱偵僇僗儁乕僗旕埶懚揑側嵶朎巰偑娭梌偡傞偙偲偑傢偐偭偰偒偨丅偦偙偱丄JNK宱楬偑偙傟傜偺夁掱偵娭梌偡傞偐偳偆偐傪娷傔丄JNK埶懚揑嵶朎巰偺堄媊偵偮偄偰専摙偟偰偄傞丅
仺JNK偵傛傞嵶朎巰桿摫婡峔偺夝愅
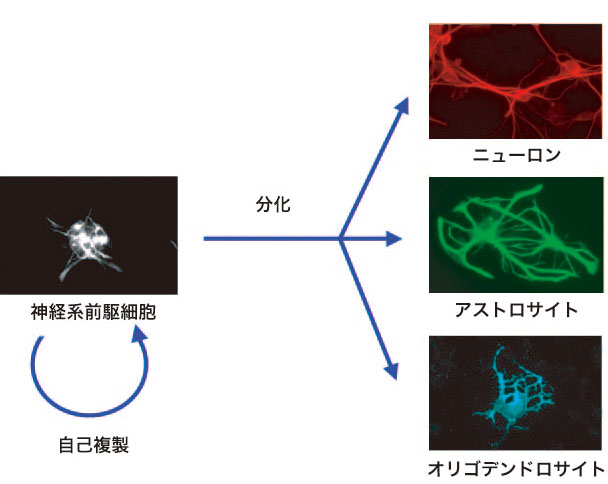
俁丏恄宱宯慜嬱嵶朎偺塣柦惂屼
丂
恄宱宯慜嬱嵶朎(neural precursor cells) 偼丄枹暘壔偱帺屓暋惢擻傪桳偟丄條乆側僯儏乕儘儞傗僌儕傾(傾僗僩儘僒僀僩丄僆儕僑僨儞僪儘僒僀僩乯偵暘壔偡傞擻椡傪堐帩偟偰偄傞嵶朎偱偁傞丅戝擼旂幙偺恄宱宯慜嬱嵶朎偼丄傑偢戀惗憗婜偵偁傞掱搙帺屓暋惢乮枹暘壔側傑傑憹怋乯偟丄偦偺屻堦晹偑戀惗屻婜偵僯儏乕儘儞傊偲暘壔偟偰丄堦晹偑弌惗屻偵僌儕傾傊偲暘壔偡傞偙偲偑抦傜傟偰偄傞丅偙偺帺屓暋惢偐傜暘壔傊偲塣柦曄姺偡傞僞僀儈儞僌偺挷愡偼丄揔惓側悢偺僯儏乕儘儞偲僌儕傾嵶朎傪嶻傒弌偡偺偵旕忢偵廳梫偱偁傞偑丄偦偺惂屼儊僇僯僘儉偼杦偳柧傜偐偵側偭偰偄側偄丅変乆偼丄戀惗憗婜偺戝擼恄宱宯慜嬱嵶朎偵偍偄偰丄Notch僔僌僫儖偲FGF僔僌僫儖偑STAT3傪妶惈壔偟丄STAT3偑偙傟傜偺僔僌僫儖偵傛傞恄宱宯慜嬱嵶朎偺帺屓暋惢擻偺懀恑偵昁恵偺栶妱傪壥偨偡偙偲傪柧傜偐偵偟偨丅偟偐偟丄摨偠STAT3偼戀惗屻婜偺戝擼恄宱宯慜嬱嵶朎偵偍偄偰偼傾僗僩儘僒僀僩暘壔傪懀恑偡傞偙偲偑抦傜傟偰偄傞丅傑偨変乆偼丄憗婜偺戝擼旂幙恄宱宯慜嬱嵶朎偱偼帺屓暋惢傪懀恑偡傞Wnt僔僌僫儖偑丄屻婜偱偼帺屓暋惢傪傓偟傠慾奞偟僯儏乕儘儞暘壔傪懀恑偡傞偙偲傪尒弌偟偨丅峏偵変乆偼丄憗婜偺戝擼恄宱宯慜嬱嵶朎偱偼帺屓暋惢偺懀恑偵摥偔偲峫偊傜傟偰偄傞PDK1-Akt宱楬偑丄傗偼傝屻婜偱偼摿掕偺僯儏乕儘儞暘壔偵娭梌偡傞偙偲傕柧傜偐偵偟偨丅偦偙偱尰嵼摨偠応強偺恄宱宯慜嬱嵶朎偑丄帪婜偩偗偺堘偄偱側偤堎側傞僔僌僫儖墳摎惈傪帵偡偺偐傪専摙偟偰偄傞丅偙偺栤戣偼丄帺屓暋惢偐傜暘壔偵堏峴偡傞僞僀儈儞僌傪寛傔傞儊僇僯僘儉偺棟夝偵捈寢偡傞偲峫偊偰偍傝丄峏偵偙偺傛偆側恄宱宯慜嬱嵶朎偺惈幙偺棟夝偑彨棃偺嵞惗堛椕偺婎斦偲側傞偲峫偊偰偄傞丅
丂恄宱曄惉幘姵偱幐傢傟偨僯儏乕儘儞傪丄恄宱宯慜嬱嵶朎傪梡偄偰曗偆偲偄偆嵞惗堛椕偺帋傒偑巒傑偭偰偄傞丅偟偐偟側偑傜丄戝検偵昁梫側恄宱宯慜嬱嵶朎偺嫙媼尮傪摼傞堊偵丄in
vitro偱恄宱宯慜嬱嵶朎傪攟梴偟憹怋偝偣偰傕丄暘楐夞悢傪廳偹傞偵敽偭偰帺屓暋惢擻傪幐偄丄摿掕偺暘壔宍幙偵僐儈僢僩偟偰偟傑偆偙偲偱丄昁梫側僯儏乕儘儞偑摼傜傟側偄偲偄偆偺偑丄尰忬偵偍偗傞旕忢偵戝偒側栤戣偱偁傞丅偙偺揰偵懳張丒崕暈偡傞偨傔偵傕丄乽暘楐夞悢傪廳偹傞偙偲偱壗偑婲偒偰偄傞偺偐乿傪棟夝偡傞偙偲偑廳梫側壽戣偱偁傞丅
弶婜偺恄宱宯敪惗偵偍偄偰旕忢偵懡偔偺嵶朎偑巰偸 偙偲偑抦傜傟偰偄傞丅偙偺嵶朎巰傪椺偊偽僇僗儁乕僗俋偺堚揱巕攋夡偵傛偭偰梷惂偡傞偲丄摿偵枹暘壔側恄宱宯慜嬱嵶朎偑懡偔側傝偡偓偰擼偑堎忢偵旍戝偡傞丅偙偺偙偲偼丄恄宱宯慜嬱嵶朎偺巰偑惓忢側恄宱宯偺敪惗偵昁恵偱偁傞偙偲傪抂揑偵帵偟偰偄傞偲尵偊傞丅
変乆偼丄恄宱宯慜嬱嵶朎偺惗懚傪Notch僔僌僫儖偑懀恑偡傞偙偲乮嫽枴怺偄偙偲偵丄枹暘壔惈堐帩偲偼暿偺宱楬偱両乯傪尒弌偟偰偄傞丅丄